General
Projects
Development of Neuronal Morphology

In collaboration with Dr Arjen van Ooyen (Center for Neurogenomics and Cognitive Research, VU University, Amsterdam), I am investigating biophysically realistic mathematical models of neurite outgrowth. A particular emphasis is on the growth of dendritic trees for different neuronal types. We are also developing appropriate numerical techniques for the computer simulation of these models. A review of this and related work can be found in (Graham and van Ooyen, 2006).
- Neurite Elongation
- Simple Models of Neurite Branching
- Complex Models of Elongation and Branching
- Compartmental Modelling Techniques
- Funding
- Key References
- Related References
Neurite Elongation [top]
We have recently developed a continuum (PDE) model of tubulin-driven neurite elongation that allows determination of tubulin concentration and transport along the length of the growing neurite (McLean et al., 2004; Graham et al., 2006).
Analysis of this model by McLean (McLean et al., 2004a, 2005) has revealed that growth to a steady-state length can occur in three different growth regimes, determined by the relative proportions of construction, due to the synthesis and active transport of tubulin and its assembly onto microtubules, and dissipation, due to tubulin diffusion, degradation and its disassembly from microtubules.
The dynamics of outgrowth have been explored using analysis (McLean and Graham, 2006) and a numerical solution of the model (Graham et al., 2006; McLean et al., 2004b). Outgrowth is stable and is usually quite damped. Oscillations in length can occur when growth is affected by both the active transport and diffusion of tubulin. Autoregulation of tubulin production is able to damp out these oscillations.
Matlab scripts implementing the numerical solution of the model are available for download.
Simple Models of Neurite Branching [top]
Using simple models based on the transport of tubulin in the growing neurite we have examined transport-limiting effects on competitive outgrowth (van Ooyen, 2001) and branching (Graham and van Ooyen, 2004; van Pelt et al., 2003).
These models are defined as sets of coupled ordinary differential equations (ODEs) that each describe the transport of the elongation or branch-determining substance along one unbranched segement of the developing tree. The concentration of the substance at the tip of each terminal segment determines either the rate of elongation (van Ooyen, 2001) or the probability that the terminal will split to form two daughter branches (Graham and van Ooyen, 2004).
The branching model shows that modulations in branching probability during outgrowth, identified by statistical models of dendrite formation (van Pelt and Uylings, 1999), can arise from variations in the availability of a branch-determining substance at each terminal due to the diffusion and active transport of that substance from its site of production in the cell body. Such a substance can plausibly be identified as tubulin, or possibly a microtubule-associated protein, such as MAP2, that influences the stability of microtubule bundles in dendrites.
Here is a movie of a developing, branching neurite from our model (AVI format, 265KB).
A Java-based simulator (with source code) for this branching model is available for download.
Complex Models of Elongation and Branching [top]
PhD student Gregor Kiddie (on a project studentship provided by an EPSRC grant), has been formulating a much more sophisticated model, based on the work of Hely (Hely et al. 2001), that describes both the elongation rate and the branching rate of a neurite tip explicitly as functions of tubulin and MAP2 concentrations at the tip (Kiddie et al. 2005). The aim is to explore whether both the segment length distributions and branching structures found in real dendrites may arise from the constraints on cytoskeleton construction.
Further details are described in this poster, presented at FENS in Lisbon, 2004. Below is an image from Hely's original model showing some time sequences of neurite outgrowth for different types of neurite.
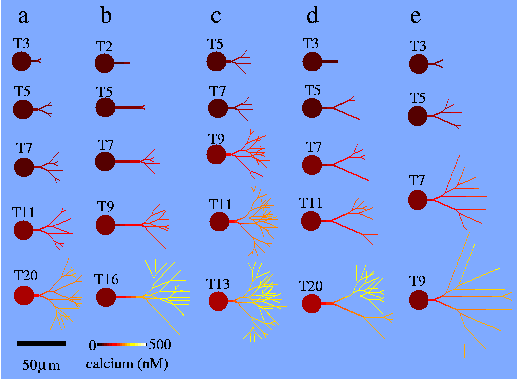
Compartmental Modelling Techniques [top]
Computer simulation of these models requires numerical techniques that can cope with the changing morphology of the structure being simulated ie the growing neurite. We are currently using two methods.
One method uses a spatial discretization with a fixed number of grid points (or "compartments"), with the grid points moving further apart as a neurite elongates. Full details can be in found in (Graham et al., 2006) and these Matlab scripts.
The other method uses "compartments" of a fixed length, with new compartments being added as required to increase a neurite's length and create new branches. Initial work has established basic strategies for the addition and deletion of compartments while preserving accurate calculation of quantities of interest (Graham and van Ooyen, 2001). An implementation is provided in this Java-based simulator.
In a review paper (Graham and van Ooyen, 2006) we discuss possibilities for a complete software package for simulating the development of neuronal morphology.
Funding [top]
- 2002-2005
- EPSRC Grant GR/R89769/01, Ł64,811, to B. Graham with project partner A. van Ooyen. This grant provided a project studentship to G. Kiddie. Please read our final narrative report.
Key References [top]
Graham, B. and van Ooyen, A. (2006). Mathematical modelling and numerical simulation of the morphological development of neurons. BMC Neuroscience, 7 (Suppl 1): S9.
Graham, B., Lauchlan, K., and McLean, D. (2006). Dynamics of outgrowth in a continuum model of neurite elongation. J. Comput. Neurosci., 20:43-60.
Graham B.P. and van Ooyen, A. (2004). Transport limited effects in a model of dendritic branching. J. Theor. Biol., 230:421-432.
Graham, B. and van Ooyen, A. (2001).
Compartmental models of growing neurites.
Neurocomputing, 38-40:31-36.
Hely, T., Graham, B., and van Ooyen, A. (2001).
A computational model of dendrite elongation and branching based on
MAP2 phosphorylation.
J. Theor. Biol., 210:375-384.
Kiddie, G., McLean, D., van Ooyen, A. and Graham, B. (2005). Biologically plausible models of neurite outgrowth. Progress in Brain Research, 147: 67-80.
McLean, D.R. and Graham, B.P. (2006). Stability in a mathematical model of neurite elongation. Mathematical Medicine and Biology, 23:101-117.
McLean, D., Lauchlan, K., and Graham, B. (2005). On the existence of steady solutions in a moving boundary model of neurite morphogenesis with cellular autoregulation. WSEAS Transactions on Biology and Biomedicine, 1:98-105.
McLean, D.R. and Graham, B.P. (2004). Mathematical formulation and analysis of a continuum model for tubulin-driven neurite elongation. Proc. R. Soc. Lond. A, 460:2437-2456.
McLean, D.R., van Ooyen, A. and Graham, B.P. (2004). Continuum model for tubulin-driven neurite elongation. Neurocomputing, 58-60:511-516.
van Ooyen, A., Graham, B., and Ramakers, G. (2001).
Competition for tubulin between growing neurites during development.
Neurocomputing, 38-40:73-78.
van Pelt, J., Graham, B., and Uylings, H. (2003).
Formation of dendritic branching patterns.
In van Ooyen, A., editor, Modeling Neuronal Development,
chapter 4, MIT Press, Cambridge, MA.
Related References [top]